 Вход
ВходСтруктура и виды РНК. Роль РНК в процессе реализации наследственной информации. Структура и функции рнк Структура молекулы рнк представляет собой
Первичная структура РНК – порядок чередования рибонуклеозидмонофосфатов в полинуклеотидной цепи. В РНК, как и в ДНК, нуклеотиды связаны между собой 3",5"-фосфодиэфирными связями. Концы полинуклеотидных цепей РНК неодинаковы. На одном конце находится фосфорилированная ОН-группа 5"-углеродного атома, на другом конце – ОН-группа 3"-углеродного атома рибозы, поэтому концы называют 5"- и 3"-концами цепи РНК.
Вторичная структура РНК
Молекула рибонуклеиновой кислоты построена из одной полинуклеотидной цепи. Отдельные участки цепи РНК образуют спирализованные петли – «шпильки», за счёт водородных связей между комплементарными азотистыми основаниями A-U и G-C. Участки цепи РНК в таких спиральных структурах антипараллельны, но не всегда полностью комплементарны, в них встречаются неспаренные нуклеотидные остатки или даже одноцепочечные петли, не вписывающиеся в двойную спираль. Наличие спирализованных участков характерно для всех типов РНК.
Третичная структура РНК
Одноцепочечные РНК характеризуются компактной и упорядоченной третичной структурой, возникающей путём взаимодействия спирализованных элементов вторичной структуры. Так, возможно образование дополнительных водородных связей между нуклеотидными остатками, достаточно удалёнными друг от друга, или связей между ОН-группами остатков рибозы и основаниями. Третичная структура РНК стабилизирована ионами двухвалентных металлов, например ионами Mg 2+ , связывающимися не только с фосфатными группами, но и с основаниями.
Основные типы рнк
В цитоплазме клеток присутствуют 3 типа рибонуклеиновых кислот – транспортные РНК (тРНК), матричные РНК (мРНК) и рибосомальные РНК (рРНК). Они различаются по первичной структуре, молекулярной массе, конформации, продолжительности жизни и, самое главное, по функциональной активности.
http :// www . biochemistry . ru / biohimija _ severina / B 5873 Part 25-141. html
Методы определения первичной и вторичной структуры нуклеиновых кислот
Секвенирование – это общее название методов, которые позволяют установить последовательность нуклеотидов в молекуле ДНК. В настоящее время нет ни одного метода секвенирования, который бы работал для молекулы ДНК целиком; все они устроены так: сначала готовится большое число небольших участков ДНК (клонируется молекула ДНК многократно и «разрезается» её в случайных местах), а потом читается каждый участок по отдельности.
Клонирование происходит либо просто выращиванием клеток в чашке Петри, либо (в случаях, когда это было бы слишком медленно или по каким-то причинам не получилось бы) при помощи так называемой полимеразной цепной реакции. В кратком и неточном изложении работает она примерно так: сначала ДНК денатурируют, т.е. разрушают водородные связи, получая отдельные нити. Затем к ДНК присоединяют так называемые праймеры; это короткие участки ДНК, к которым может присоединиться ДНК-полимераза – соединение, которое, собственно, и занимается копированием (репликацией) нити ДНК . На следующем этапе полимераза копирует ДНК, после чего процесс можно повторять: после новой денатурации отдельных нитей будет уже вдвое больше, на третьем цикле – вчетверо, и так далее.
Все эти эффекты достигаются в основном с помощью изменений температуры смеси из ДНК, праймеров и полимеразы; для наших целей важно, что это достаточно точный процесс, и ошибки в нём редки, а на выходе получается большое число копий участков одной и той же ДНК. Разные методы секвенирования отличаются друг от друга не методами клонирования, а тем, как потом прочесть получившийся «суп» из многочисленных копий одной и той же ДНК.
Метод ДНК-ДНК гибридизации основан на том факте, что стабильность ДНК-ДНК дуплексов при определенной температуре зависит от числа нуклеотидов образующих комплементарные пары. Очевидно, что число комплементарных нуклеотидов в дуплексе где обе нити происходят из одной и той же молекулы ДНК (т.е. в гомодуплексах) равно 100%. Если же обе нити имеют разное происхождение (гетеродуплекс), то, в зависимости от числа произошедших мутаций, число комплементарных пар будет меньше 100%. Соответсвенно гетеродуплексы должны распадаться (плавится) при более низкой температуре, чем гомодуплексы. Причем, чем ниже температура плавления, тем больше различия в двух последовательностях. Температурная стабильность гибридной ДНК определяется температурой при которой 50% гибридной ДНК диссоциировалось в одноцепочечную форму. Затем эта температура сравнивается со средней температурой 50%-го плавления гомодуплексов обоих типов последовательностей участвующих в образовании гетеродуплекса, эта температура обычно обозначается Tm. Разница между медианной температурой плавления гетеро- и гомодуплексов обозначается как dTm. Показана линейная зависимость dTm от числа неспаренных оснований ( Britten et. al., 1974 ): p=cdTm. Константа c обычно определяется условиями проведения эксперимента и обычно варьирует от 0.01 до 0.015. Определение dTm требует большого числа повторений, т.к. велика экспериментальная ошибка.
Основным свойством ДНК является ее способность к репликации.
http :// postnauka . ru / longreads /468
1.9. Репликация ДНК, транскрипция, трансляция, обратная транскрипция. Амплификация ДНК. Биосинтез белка, аминокислотный код. Организация генов, строение генов у про- и эукариот, понятие о клонировании.
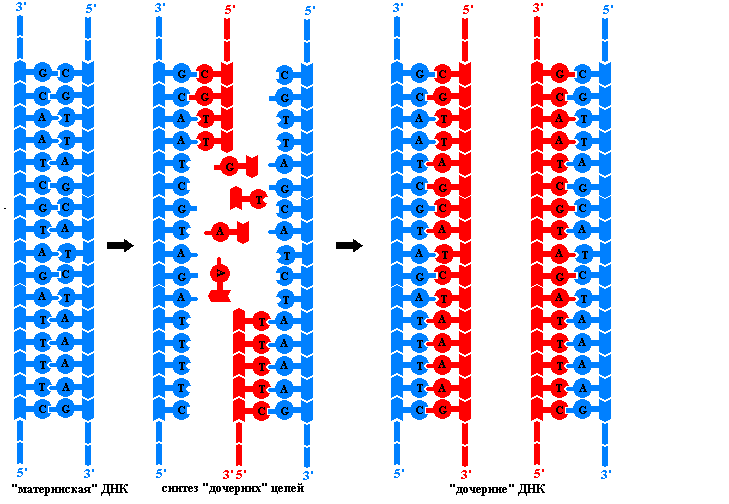
Репликация – это процесс самоудвоения молекул ДНК, происходящий под контролем ферментов. Репликация осуществляется перед каждым делением ядра. Начинается она с того, что спираль ДНК временно раскручивается под действием фермента ДНК-полимеразы. На каждой из цепей, образовавшихся после разрыва водородных связей, по принципу комплементарности синтезируется дочерняя цепь ДНК. Материалом для синтеза служат свободные нуклеотиды, которые есть в ядре.
Схема репликации ДНК
Таким образом, каждая полинуклеотидная цепь выполняет роль матрицы для новой комплементарной цепи (поэтому процесс удвоения молекул ДНК относится к реакциям матричного синтеза). В результате получается две молекулы ДНК, у каждой из которых одна цепь остается от родительской молекулы (половина), а другая – вновь синтезированная. Причем одна новая цепь синтезируются сплошной, а вторая – сначала в виде коротких фрагментов, которые затем сшиваются в длинную цепь специальным ферментом – ДНК-лигазой. В результате репликации две новые молекулы ДНК представляют собой точную копию исходной молекулы.
Биологический смысл репликации заключается в точной передаче наследственной информации от материнской клетки к дочерним, что и происходит при делении соматических клеток.
http :// sbio . info / page . php ? id =11
Литература:
1) Н. Грин, У. Стаут, Д. Тейлор – Биология.
2) З.А. Шабарова и А.А. богданов – Химия нуклеиновых кислот и их полимеров.
3) А.П. Пехов – Биология и общая гинетика.
4) А. Микельсон – Химия нуклеозидов и нуклеотидов.
5) З. Гауптман, Ю. Грефе, Х. Ремане – Органическая химия
Транскри́пция – это процесс синтеза РНК с использованием ДНК в качестве матрицы, происходящий во всех живых клетках. Другими словами, это перенос генетической информации с ДНК на РНК.
Транскрипция катализируется ферментом ДНК-зависимой РНК-полимеразой. Процесс синтеза РНК протекает в направлении от 5"- к 3"- концу, то есть по матричной цепи ДНК РНК-полимераза движется в направлении 3"- 5".
Транскрипция состоит из стадий инициации, элонгации и терминации. Единицей транскрипции является транскриптон, фрагмент молекулы ДНК, состоящий из промотора, транскрибируемой части и терминатора.
Инициация транскрипции – это сложный процесс, зависящий от последовательности ДНК вблизи транскрибируемой последовательности (а у эукариот также и от более далеких участков генома - энхансеров и сайленсеров ) и от наличия или отсутствия различных белковых факторов .
Элонгация транскрипции
Момент перехода РНК-полимеразы от инициации транскрипции к элонгации точно не определен. Три основных биохимических события характеризуют этот переход в случае РНК-полимеразы кишечной палочки : отделение сигма-фактора, первая транслокация молекулы фермента вдоль матрицы и сильная стабилизация транскрипционного комплекса, который кроме РНК-полимеразы включает растущую цепь РНК и транскрибируемую ДНК. Эти же явления характерны и для РНК-полимераз эукариот. Переход от инициации к элонгации сопровождается разрывом связей между ферментом, промотором , факторами инициации транскрипции, а в ряде случаев – переходом РНК-полимеразы в состояние компетентности в отношении элонгации. Фаза элонгации заканчивается после освобождения растущего транскрипта и диссоциации фермента от матрицы (терминация).
На стадии элонгации в ДНК расплетено примерно 18 пар нуклеотидов . Примерно 12 нуклеотидов матричной нити ДНК образует гибридную спираль с растущим концом цепи РНК. По мере движения РНК-полимеразы по матрице впереди нее происходит расплетание, а позади – восстановление двойной спирали ДНК. Одновременно освобождается очередное звено растущей цепи РНК из комплекса с матрицей и РНК-полимеразой. Эти перемещения должны сопровождаться относительным вращением РНК-полимеразы и ДНК.
73. Строение рнк. Виды рнк. Их роль в метаболизме.
Рибонуклеиновая кислота (РНК) – это однонитевой биополимер, в качестве мономеров которого выступают нуклеотиды.
Матрицей для синтеза новых молекул РНК являются молекулы дезоксирибонуклеиновой кислоты (транскрипция РНК). Хотя в ряде случаев возможен и обратный процесс (образование новых ДНК на матрице РНК в ходе репликации некоторых вирусов). Также основой для биосинтеза РНК могут быть другие молекулы рибонуклеиновой кислоты (репликация РНК). В транскрипции РНК, происходящей в ядре клетки, участвует целый ряд ферментов, наиболее значимым из которых является РНК-полимераза.
Структура РНК.
Молекула имеет однонитевое строение. Полимер. В результате взаимодействия нуклеотидов друг с другом молекула РНК приобретает вторичную структуру, различной формы (спираль, глобула и т.д.). Мономером РНК является нуклеотид (молекула, в состав которой входит азотистое основание, остаток фосфорной кислоты и сахар (пептоза)). РНК напоминает по своему строению одну цепь ДНК. Нуклеотиды, входящие в состав РНК: гуанин, аденин, цитозин, урацил. Аденин и гуанин относятся к пуриновым основаниям, цитозин и урацил к пиримидиновым. В отличие от молекулы ДНК, в качестве углеводного компонента рибонуклеиновой кислоты выступает не дезоксирибоза, а рибоза. Вторым существенным отличием в химическом строении РНК от ДНК является отсутствие в молекуле рибонуклеиновой кислоты такого нуклеотида как тимин. В РНК он заменён на урацил.
Функции РНК различаются в зависимости от вида рибонуклеиновый кислоты.
1) Информационная РНК (и-РНК).
Иногда данный биополимер называют матричной РНК (м-РНК). Данный вид РНК располагается как в ядре, так и в цитоплазме клетки. Основное назначение – перенос информации о строении белка от дезоксирибонуклеиновой кислоты к рибосомам, где и происходит сбор белковой молекулы. Относительно небольшая популяция молекул РНК, составляющая менее 1% от всех молекул.
2) Рибосомная РНК (р-РНК).
Самый распространенный вид РНК (около 90% от всех молекул данного вида в клетке). Р-РНК расположена в рибосомах и является матрицей для синтеза белковых молекул. Имеет наибольшие, по сравнению с другими видами РНК, размеры. Молекулярная масса может достигать 1,5 миллионов кДальтон и более.
3) Транспортная РНК (т-РНК).
Расположена, преимущественно, в цитоплазме клетки. Основное назначение- осуществление транспорта (переноса) аминокислот к месту синтеза белка (в рибосомы). Транспортная РНК составляет до 10% от всех молекул РНК, располагающихся в клетке. Имеет наименьше, по сравнению с другими РНК- молекулами, размеры (до 100 нуклеотидов).
4) Минорные (малые) РНК.
Это молекулы РНК, чаще всего с небольшой молекулярной массой, располагающиеся в различных участках клетки (мембране, цитоплазме, органеллах, ядре и т.д.). Их роль до конца не изучена. Доказано, что они могут помогать созреванию рибосомной РНК, участвуют в переносе белков через мембрану клетки, способствуют редупликации молекул ДНК и т.д.
5) Рибозимы.
Недавно выявленный вид РНК, принимающие активное участие в ферментативных процессах клетки в качестве фермента (катализатора).
6) Вирусные РНК.
Любой вирус может содержать только один вид нуклеиновой кислоты: либо ДНК либо РНК. Соответственно, вирусы, имеющие в своём составе молекулу РНК, получили название РНК-содержащие. При попадании в клетку вируса данного типа может происходить процесс обратной транскрипции (образование новых ДНК на базе РНК), и уже вновь образовавшаяся ДНК вируса встраивается в геном клетки и обеспечивает существование, а также размножение возбудителя. Вторым вариантом сценария является образование комплиментарной РНК на матрице поступившей вирусной РНК. В этом случае, образование новых вирусных белков, жизнедеятельность и размножение вируса происходит без участия дезоксирибонуклеиновой кислоты только на основании генетической информации, записанной на вирусной-РНК.
РНК,как и ДНК, представляет собой полинуклеотид. Структура нуклеотидов РНК с таковой ДНК, но имеются следующие отличия:
- Вместо дезоксирибозы в состав нуклеотидов РНК входит пятиуглеродный сахар- рибоза;
- Вместо азотистого основания тимина- урацил;
- Молекула РНК обычно представлена одной цепочкой (у некоторых вирусов- двумя);
В клетках существуют три типа РНК: информационная,транспортная и рибосомальная.
Инфармационная РНК (и-РНК) представляет собой копию определённого участка ДНК и выполняет роль переносчика генетической информации от ДНК к месту синтеза белка (рибосомы) и непосредственно участвует в сборке его молекул.
Транспортные РНК (т-РНК)переносят аминокислоты из цитоплазмы в рибосомы.
Рибосомальная РНК (р-РНК) входит в состав рибосом. Считают, что р-РНК обеспечивает определённое пространственное взаиморасположение и-РНК и т-РНК.
Роль РНК в процессе реализации наследственной информации.
Наследственная информация, записанная с помощью генетического кода, хранится в молекулах ДНК и размножается для того, чтобы обеспечить вновь образуемые клетки необходимыми «инструкциями» для их нормального развития и функционирования. Вместе с тем непосредственного участия в жизнеобеспечении клеток ДНК не принимает. Роль посредника, функцией которого является перевод наследственной информации, сохраняемой в ДНК, в рабочую форму, играют рибонуклеиновые кислоты - РНК.
В отличие от молекул ДНК рибонуклеиновые кислоты представлены одной полинуклеотидной цепью, которая состоит из четырех разновидностей нуклеотидов, содержащих сахар, рибозу, фосфат и одно из четырех азотистых оснований - аденин, гуанин, урацил или цитозин. РНК синтезируется на молекулах ДНК при помощи ферментов РНК-полимераз с соблюдением принципа комплементарности и антипараллельности, причем аденину ДНК в РНК комплементарен урацил. Все многообразие РНК, действующих в клетке, можно разделить на три основных вида: мРНК, тРНК, рРНК.
По химической организации материала наследственности и изменчивости эукариотические и прокариотические клетки принципиально не отличаются друг от друга. Генетический материал у них представлен ДНК. Общим для них является и принцип записи генетической информации, а также генетический код. Одни и те же аминокислоты шифруются у про- и эукариот одинаковыми кодонами. Принципиально одинаковым образом у названных типов клеток осуществляется и использование наследственной информации, хранящейся в ДНК. Сначала она транскрибируется в нуклеотидную последовательность молекулы мРНК, а затем транслируется в аминокислотную последовательность пептида на рибосомах с участием тРНК. Однако некоторые особенности организации наследственного материала, отличающие эукариотические клетки от прокариотических, обусловливают различия в использовании их генетической информации.
Наследственный материал прокариотической клетки содержится главным образом в единственной кольцевой молекуле ДНК. Она располагается непосредственно в цитоплазме клетки, где также находятся необходимые для экспрессии генов тРНК и ферменты, часть из которых заключена в рибосомах. Гены прокариот состоят целиком из кодирующих нуклеотидных последовательностей, реализующихся в ходе синтеза белков, тРНК или рРНК.
Наследственный материал эукариот больше по объему, чем у прокариот. Он расположен в основном в особых ядерных структурах -хромосомах , которые отделены от цитоплазмы ядерной оболочкой. Необходимый для синтеза белков аппарат, состоящий из рибосом, тРНК, набора аминокислот и ферментов, находится в цитоплазме клетки.
Значительные отличия имеются в молекулярной организации генов эукариотической клетки. В большинстве из них кодирующие последовательности экзоны прерываются интронными участками, которые не используются при синтезе т-РНК, р-РНК или пептидов. Количество таких участков варьирует в разных генах.. Эти участки удаляются из первично-транскрибируемой РНК, в связи с чем использование генетической информации в эукариотической клетке происходит несколько иначе. В прокариотической клетке, где наследственный материал и аппарат биосинтеза белка пространственно не разобщены, транскрипция и трансляция происходят почти одновременно. В эукариотической клетке эти два этапа не только пространственно отделены ядерной оболочкой, но и во времени их разделяют процессы созревания м-РНК, из которой должны быть удалены неинформативные последовательности.
Кроме указанных различий на каждом этапе экспрессии генетической информации можно отметить некоторые особенности течения этих процессов у про- и эукариот.
В отличие от ДНК, молекула РНК состоит из одной полинуклеотидной цепи, которая спирализована сама на себя, т.е. образует всевозможные «петли» и «шпильки» за счет взаимодействий комплементарных азотистых оснований (вторичная структура). У некоторых вирусов встречаются двуцепочечные РНК, которые несут генетическую информацию аналогично ДНК.
Существуют:
1 – матричные РНК (мРНК);
2 – рибосомные РНК (рРНК);
3 – транспортные РНК (тРНК).
Рибосомные РНК. На долю рРНК приходится 80-90% клеточной РНК. Локализованы в рибосомах, в комплексе с рибосомными белками. Рибосомы состоят из двух частей и представляют собой нуклеопротеины, состоящие из рРНК и белка в соотношении 1:1 (для эукариот) и 2:1 (для прокариот).
Биологическая роль рРНК – являются структурной основой рибосом, взаимодействует с мРНК и тРНК в процессе биосинтеза белка, принимает участие в процессе сборки полипептидной цепи.
У эукариот обнаружено 4 типа рРНК с различным коэф. седиментации: 18S(в малой части рибосомы), а 28S, 5,8S и 5S (сведбергов) – в большой части рибосомы.. Они различаются молекулярной массой (35 000-1 600 000) и локализацией в рибосомах.
Вторичная структура рРНК характеризуется спирализацией цепи самой на себя, третичная – ее компактной укладкой.
Матричные РНК. Матричная РНК составляет 2-3% от всей клеточной РНК, синтезируется мРНК в ядре клетки на матрице ДНК (процесс транскрипции), переписывая с нее генетическую информацию по принципу комплементарности.
ДНК -А-Т-Г-Ц-
ДНК -Т-А-Ц-Г-
мРНК -А-У-Г-Ц-
Затем мРНК поступают в цитоплазму, соединяются с рибосомой и выполняют роль матрицы для биосинтеза белка. Каждой аминокислоте соответствует в мРНК определенная тройка (триплет) нуклеотидов, называемая кодоном этой аминокислоты. Последовательность кодонов в цепи мРНК определяет последовательность аминокислот в белке. Всего может быть 64 кодона. Из них 61 кодон кодирует аминокислоты, а 3 кодона – кодоны терминаторы (терминирующие), которые обозначают окончание белкового синтеза. Существуют также инициирующие кодоны, которые соответствуют первой аминокислоте в белке и чаще всего соответствуют аминокислоте метионину.
Поскольку мРНК несет наследственную информацию о первичной структуре белка, нередко ее называют информационной РНК (иРНК). Каждый отдельный белок, синтезируемый в клетке, кодируется определенной «своей» мРНК или ее участком. мРНК образует несколько двуспиральных «шпилек», на концах которых располагаются знаки (например, ААУААА) инициации (начала синтеза белка) и терминации (окончания синтеза белка).
Т.о. информация о строении белка закодирована в ДНК с помощью генетического кода, который является линейным, непрерывным, триплетным, выражденным. Он является универсальным.
Молекулярный вес мРНК варьирует в широких пределах от 35 000 до нескольких млн. мРНК ранее считались короткоживущими РНК. Для микроорганизмов время жизни мРНК несколько секунд или минут. Но для эукариот – оно может составлять от нескольких часов до нескольких недель.
Транспортная РНК. Составляют 10-20% клеточной РНК.
Функции тРНК:
1 - связывают аминокислоты и транспортируют их в рибосому, где происходит синтез белка;
2 – кодируют аминокислоты;
3 – Расшифровывают генетический код.
Каждая тРНК может переносить только 1 строго определенную аминокислоту.
тРНК именуются по названию аминокислот. Например, аланиновая тРНК. тРНК, связывающие одну и ту же аминокислоту, называют изоакцепторными и нумеруют: тРНК 1 вал, тРНК 2 вал и т.д.
Вторичная структура всех тРНК имеет форму «клеверного листа». В его составе различают:
1. акцепторный стебель – к нему присоединяется аминокислота.
2. Псевдоуридиловая петля – используется для связи тРНК с рибосомой.
3. Дополнительная петля – назначение неизвестно.
4. Антикодоновая петля – содержит антикодон (триплет нуклеиновых остатков, которые комплементарны кодону мРНК, с его помощью тРНК соединяется с мРНК);
5. Дигидроуридиновая петля – обеспечивает связывание тРНК со специфическим ферментом (аминоацил-тРНК-синтетазой), который соединяет аминокислоту с тРНК.
Стабилизируется вторичная структура водородными связями между комплементарными основаниями.
Третичная структура тРНК имеет неправильную Г-образную форму. стабилизирована водородными и др. связями.
Молекула РНК также полимер, мономерами которого является рибонуклеотиды, РНК представляет собой одноцепочную молекулу. Она построена таким же образом, как и одна из цепей ДНК. Нуклеотиды РНК похожи на нуклеотиды ДНК, хотя и не тождественны им. Их тоже четыре, и они состоят из осатков азотистого основания, пентозы и фосфорной кислоты. Три азотистых основания совершенно такие же, как в ДНК: А , Г и Ц . Однако вместо Т у ДНК в РНК присутствует близкое по строение пиримидиновое основание – урацил (У ). основное различие между ДНК и РНК – это характер углевода: в нуклотидах ДНК моносахарид – дезоксирибоза, а в РНК – рибоза. Связь между нуклеотидами осуществляется, как и в ДНК, через сахар и остаток фосфорной кислоты. В отличие от ДНК, содержание которой в клетках определенных организмов постоянно, содержание РНК в них колеблется. Оно заметно выше там, где происходит интенсивный синтез.
В отношении выполняемых функций различают несколько видов РНК.
Транспортная РНК (тРНК). Молекулы тРНк самые короткие: они состоят всего из 80-100 нуклеотидов. Молекулярная масса таких частиц равна 25-30 тыс. Транспортные РНК в основном содержатся в цитоплазме клетки. Функция их состоит в переносе аминокислот в рибосомы, к месту синтеза белка. Из общего содержания РНК клетк на долю тРНК приходится около 10%.
Рибосомная РНК (рРНК). Это крупные молекулы: в их состав входит 3-5 тыс. нуклеотидов, соотвественно их молекулярная масса достигает 1-1,5 млн. Рибосомные РНК составляют существенную часть рибосомы. Из общего содержания РНК в клетке на долю рРНК приходится около 90%.
Информационная РНК (иРНК), или матричная РНК (мРНК), содержится в ядре и цитоплазме. Функция ее состоит в переносе информации о структуре белка от ДНК к месту синтеза белка в рибосомах. На долю иРНК приходится примерно 0,5-1% от общего содержания РНК клетки. Размер иРНК колеблется в широких пределах – от 100 до 10000 нуклеотидов.
Все виды РНК синтезируются на ДНК, которая служитсвоего рода матрицей.
ДНК – носитель наследсвенной нформации.
Каждый белок представлен одной или несколькими полипиптидными цепями. Участок ДНК, несущий информацию об одной полипиптидной цепи, называют геном . Совокупность молекул ДНК клетки выполняет функцию носителя генетической информации. Генетическая информация передается как от материнской клетки дочерним клеткам, так и от родителей детям. Ген является единицей генетической , или наследственной, информации.
ДНК – носитель генетической информаци в клетке – непосредственного участия в синтезе белков не принимает. В клетках эукариот молекулы ДНК содержатся в хромосомах ядра и отделены ядерной оболочкой от цитоплазмы, где происходит синтез белков. К рибосомам – местам сборки белков – высылается из ядра несущий информацию посредник, способный пройти через поры ядерной оболочки. Таким посредником является информационная РНК (иРНК). По принципу комплементарности она синтезируется на ДНК при участие фермента, называемого РНК-полимеразой .

Информационная РНК – это однонитевая молекула, и транскрипция идет с одной цепи двунитевой молекулы ДНК. Она является копией не всей молекулы ДНК, а только части ее – одного гена у эукариот или группы рядом расположенных генов, несущих информацию о структуре белков, необходимых для выполнения одной функции, у прокариот. Такую группу генов называют опероном . В начале каждого оперона находится своего рода посадочная площадка для РНК-полимеразы, называемая промотором .это специфическая последовательность нуклеотидов ДНК, которую фермент «узнает» благодаря химическому сродству. Только присоединившись к промотору, РНК-полиммераза способна начать интез РНК. Доядя до конца оперона, фермент встречает сигнал (в виде определенной последоватльности нуклеотидов), означающий конец считывания. Готовая иРНК отходит от ДНК и направляется к месту синтеза белков.
В процессе транскрипции можно выделить четыре стадии: 1) связывание РНК -полимеразы с промотором; 2) инициация – начало синтеза. Оназаключается в образовании первой фосфодиэфирной связи между АТФ или ГТФ и вторым нуклеотидом синтезирующейся молекулы РНК; 3) элонгация – рост цепи РНК; т.е. последовательное присоединение нуклеотидов друг к другу в том порядке, в котором стоят комплементарные им нуклеотиды в транскрибируемой нити ДНК. Скорость элонгации 50 нуклеотидов в секунду; 4) терминация – завершение синтеза РНК.
Пройдя через поры ядерной оболочки, иРНК направляется к рибосомам, где осуществляется расшифровка генетической информации – перевод ее с «языка» нуклеотидов на «язык» аминокислот. Синтез полипептидных цепей по матрице иРНК, происходящий в рибосомах, называют трансляцией (лат. translation – перевод).
Аминокислоты, из котрых синтезируютсябелки, доставляются к рибосомам с помощью специальных РНК, называемых транспортными (тРНК). В клетке имеется столько же разных тРНК, сколько кодонов, шифрующих аминокислоты. На вершине «листа» каждой тРНК имеется последовательность трех нуклеотидов, комплементарных нуклеотидам кодона в иРНК. Ее называют антикодоном. Специальный фермент – кодаза – опознает тРНК и присоединяет к «черешку листа» аминокислоту – только ту, которая кодируется триплетом, комплементарным антикодону. На образование ковалентной связи между тРНК и «своей» аминокислотой затрачивается энергия одной молекулы АТФ.
 Для
того чтобы аминокислота включилась в
полипептидную цепь, она должна оторваться
от тРНК. Это становится возможным, когда
тРНК поступает на рибососму и антикодон
узнает свой кодон в иРНК. В рибосоме
имеется два участка для связывания двух
молекул тРНК. В один из этих участков,
называемый акцепторным
,
поступает тРНК с аминокислотой и
присоединяется к своему кодону (I).
Эта аминокислота присоединяет к себе
(акцептирует) растущую цепь белка (II)?
Между ними образуется пептидная связь.
тРНК, к которой теперь присоединяется
вместе с кодоном иРНК в донорный
участок рибосомы. В освободившийся
акцепторный участок приходит новая
тРНК, связанная с аминокислотой, которая
шифруется очередным кодоном (III).
Из донорного участка сюда вновь
переносится оторвавшаяся полипептидная
цепь и удлинняется еще на одно звено.
Аминокислоты в растущей цепи соединены
в той последовательности, в которой
расположены шифрующие их кодоны в иРНК.
Для
того чтобы аминокислота включилась в
полипептидную цепь, она должна оторваться
от тРНК. Это становится возможным, когда
тРНК поступает на рибососму и антикодон
узнает свой кодон в иРНК. В рибосоме
имеется два участка для связывания двух
молекул тРНК. В один из этих участков,
называемый акцепторным
,
поступает тРНК с аминокислотой и
присоединяется к своему кодону (I).
Эта аминокислота присоединяет к себе
(акцептирует) растущую цепь белка (II)?
Между ними образуется пептидная связь.
тРНК, к которой теперь присоединяется
вместе с кодоном иРНК в донорный
участок рибосомы. В освободившийся
акцепторный участок приходит новая
тРНК, связанная с аминокислотой, которая
шифруется очередным кодоном (III).
Из донорного участка сюда вновь
переносится оторвавшаяся полипептидная
цепь и удлинняется еще на одно звено.
Аминокислоты в растущей цепи соединены
в той последовательности, в которой
расположены шифрующие их кодоны в иРНК.
Когда на рибосоме оказывается один из трех триплетов (УАА, УАГ, УГА ), являющиеся «знаками препинания» между генами, ни одна тРНК не может занять место в акцепторном участке. Дело в том, что не существует антикодонов, комплементарных последовательностям нуклеотидов «знаков препинания». Оторвавшейся цепи не к чему присоединиться в акцепторном участке, и она покидает рибосому. Синтез белка завершен.
У прокариот синтез белков начинается с того, что кодон АУГ , расположенный на первом месте в копии с каждого гена, занимае в рибосоме такую позицию, что с ним взаимодействует антикодон особой тРНК, оединенной с формилментионином . Эта измененная форма аминокислоты метионина сразу попадает в донорный участок и выполняет роль заглавной буквы во фразе – с нее в бактериальной клетке начинается синтез любой полипептидной цепи. Когда триплет АУГ стоит не на первом месте, а внутри копии с гена, он кодирует аминокислоту метионин. После завершения синтеза полипептидной цепи формилметионин отщепляется от нее и в готовом белке отсуствует.
 Для
увеличения производства белков иРНК
часто проходит одновременно не по одной,
а по нескольким рибосомам. Акую структуру,
объединенную одной молекулой иРНК,
называют полисомой
.
На каждой рибосоме вэтом похожем на
нитку бус конвейере синтезируются
одинаковые белки.
Для
увеличения производства белков иРНК
часто проходит одновременно не по одной,
а по нескольким рибосомам. Акую структуру,
объединенную одной молекулой иРНК,
называют полисомой
.
На каждой рибосоме вэтом похожем на
нитку бус конвейере синтезируются
одинаковые белки.
Аминокислоты бесперебойно поставляются к рибосомам с помощью тРНК. Отдав аминокислоту, тРНК покидает рибосому и с помощью кодазы соединяется. Высокая слаженность всех «служб комбината» по производсву белов позволяет в течении нескольких секунд синтезировать полипептидные цепи, состоящие из сотен аминокислот.
Свойства генетического кода. Благодаря процессу транскрипции в клетке осуществляется передача информации от ДНК к белку
ДНК → иРНК → белок
Генетическая информация, содержащаяся в ДНК и в иРНК, заключена в последовательности расположения нуклеотидов в молекулах.
 Каким
же образом происходит перевод информации
с «языка» нуклеотидов на «язык»
аминокислот? Такой перевод осуществляется
с помощью генетического кода. Код,
или шифр
,
- это система символов для перевода
одной формы информации в другую.
Генетический
код
–это
система записи информации о
последовательности расположения
аминокислот в белках с помощью
последовательности расположения
нуклеотидов в иРНК.
Каким
же образом происходит перевод информации
с «языка» нуклеотидов на «язык»
аминокислот? Такой перевод осуществляется
с помощью генетического кода. Код,
или шифр
,
- это система символов для перевода
одной формы информации в другую.
Генетический
код
–это
система записи информации о
последовательности расположения
аминокислот в белках с помощью
последовательности расположения
нуклеотидов в иРНК.
Какими же свойствами обладает генетический код?
Код триплетен . В состав РНК входят четыре нуклеотида: А, Г, Ц, У. Если бы мы пытались обозначить одну аминокислоту одним нуклеотидом, то 16 из 20 аминокислот остались бы не зашифрованы. Двух буквенный код позволил бы зашифровать 16 аминокислот. Природа создала трехбуквенный, или триплетный, код. Это означает, что каждая из 20 аминокислот зашифрована последовательностью из трех нуклеотидов, называемой триплетом или кодоном.
Код вырожден. Это означает, что каждая аминокислота шифруется более чем одним кодоном. Исключения: метеонин и триптофан, каждая из которых кодируется одним триплетом.
Код однозначен. Каждый кодон шифрует только одну аминокислоту.
Между генами имеется «знаки препинания». В печатном тексте в конце каждой фразы стоит точка. Несколько связанных по смыслу фраз составляют абзац. На языке генетической информации таким абзацем являетсяоперон и комплементарная ему иРНК. Каждый ген в опероне прокариот или отдельный ген эукариот кодирует одну полипептидную цепочку – фразу. Так как в ряде случаев по матрице иРНК последовательно создается несколько разных полипептидных цепей, они должны быть отделены друг от друга. Для этого в генетическом годе имеются три специальных триплета – УАА, УАГ, УГА, каждый из которых обозначает прекращение синтеза одной полипептидной цепи. Таким образом, эти триплеты выполняют функцию знаков препинания. Они находятся в конце каждого гена.
Внутри гена нет «знаков препинания».
Код универсален. Генетический код един для всех живущих на Земле существ. У бактерий и грибов, пшеницы и хлопка, рыб и червей, лягушек и человека одни и те же триплеты кодируют одни и те же аминокислоты.

Принципы репликации ДНК. Преемственность генетического материала в поколениях клеток и организмов обеспечивается процессом репликации – удвоения молекул ДНК. Этот сложный процесс осуществляется комплексом нескольких ферментов и не обладающих каталитической активностью белов, необходимых для придания полинуклеотидным цепям нужной конформации. В результате репликации образуются две идентичные двойные спирали ДНК. Эти так называемые дочерние молекулы ничем не отличаются друг от друга и от исходной материнской молекулы ДНК. Репликация происходит в клетке перед делением, поэтому каждая дочерняя клетка получает точно такие же молекулы ДНК, какие имела материнская клетка. Процесс репликации основан на ряде принципов:
Только
в этом случае ДНК-полимеразы способна
двигаться по материнским нитям и
использовать их в качестве матриц для
безошибочного синтеза дочерних цепей.
Но полное раскручивание спиралей,
состоящих из многих миллионов пар
нуклеотидов, сопряжено со столь
значительным числом вращений и такими
энергетическими затратами, которые
невозможны в условиях клетки. Поэтому
репликация у эукариот начинается
одновременно в некоторых местах молекулы
ДНК. Участок между двумя точками, в
которых начинается синтез дочерних
цепей, называют репликоном
.
Он является единицей
репликации.


В каждой молекуле ДНК эукариотической клетки имеется много репликонов. В каждом репликоне можно видеть репликативную вилку – ту часть молекулы ДНК, которая под действием специальных ферментов уже расплелась. Каждая нить в вилке служит матрицей для синтеза комплементарной дочерней цепи. В ходе репликации вилка перемещается вдоль материнской молекулы, при этом расплетаются новые участки ДНК. Так как ДНК-полимеразы могут двигаться лишь в одном направлении вдоль матричных нитей, а нити ориентированы антипараллельно, то в каждой вилке одновременно ведут синтез два разных ферментативных комплекса. Причем в каждой вилке одна дочерняя (лидирующая) цепь растет непрерывно, а другая (отстающая) синтезируется отдельными фрагментами длинной в несколько нуклеотидов. Такие ферменты, названые в честь открывшего их японского ученого фрагментами Оказаки , сшиваются ДНК-лигазой, образуя непрерывную цепь. Механизм образования дочерних цепей ДНК фрагментами называют прерывистыми.
Потребность в затравке ДНК-полимераза не способна начинать синтез лидирующей цепи, ни синтез фрагментов Оказаки отстающей цепи. Она может лишь наращивать уже имеющуюся полинуклеотидную нить, последовательно присоединяя дезоксирибонуклеотиды к ее 3’-ОН концу. Откуда же берется начальный 5’-концевой участок растущей цепи ДНК? Его синтезирует на матрице ДНК особая РНК-полимераза, называемая праймазой (англ. Primer – затравка). Размер рибонуклеотидной затравки невелик (менее 20 нуклеотидов) в сравнении с размером цепи ДНК, образуемой ДНК-поимеразой. Выполнившая сво. Функци. РНК-затравка удаляется специальным ферментом, а образованная при эом брешь заделывается ДНК-полимеразой, использующей в качестве затравки 3’-ОН конец соседнего фрагмента Оказаки.
Проблема недорепликации концов линейных молекул ДНК. Удаление крайних РНК-праймеров, комплементрных 3’-концам обеих цепей линейной материнской молекулы ДНК, приводит к тому, что дочерние цепи оказываются короче 10-20 нуклеотидов. В этом и заключается проблема недорепликации концов линейных молекул.
Проблема недорепликации 3’-концов линейных молекул ДНК решается эукариотическими клетками с помощью специального фермента – теломеразы .
Теломераза является ДНК-полимеразой, достраивающей 3’-концылинейных молекул ДНК хромосом короткими повторяющимися последовательностями. Они, располагаясь друг за другом, образуют регулярную концевую структуру длинной до 10 тыс. нуклеотидов. Помимо белковой части, теломераза содержит РНК, выполняющую роль матрицы для наращивания ДНК повторами.
Схема удлинения концов молекул ДНК. Сначала происходит комплементарное связывание выступающего конца ДНК с матричным участком теломеразной РНК, затем теломераза наращивает ДНК, используя в качестве затравки ее 3’-ОН конец, а в качестве матрицы – РНК, входящую в состав фермента. Эта стадия называется элонгацией. После этого происходит транслокация, т.е. перемещение ДНК, удлиненной на один повтор, относительно фермента. Следом идет элонгация и очередная транслокация.
В результате образуются специализированные концевые структуры хромосом. Они состоят из многократно повторенных коротких последовательностей ДНК и специфических белков.